*
THE PLANT KINGDOM
Evolution versus involution
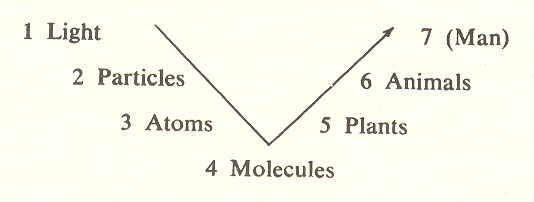
Evolution begins with the plant kingdom. Or perhaps we should say visible evolution, because the very first evolutionary activity comes earlier, with the polymers, which constitute the fifth substage of the molecular kingdom (significantly, the substage just to the right of the low point or "turn" of the arc). The polymers move against the law of entropy. And what is that?
Entropy is the tendency of the energies associated with inorganic substances to become more uniformly distributed - of stones to roll downhill, and hot objects, emitting heat, to grow cooler - so that the total energy in a given area or system gradually becomes unavailable by averaging out. This tendency, described in physics as the second law of thermodynamics, is implied in the so-called billiard ball hypothesis, which conceives of the universe as a gradually subsiding agitation of lifeless objects.
To get back to the polymers: in their self-replication they go counter to entropy; they store up energy in the substances they build. That storing up, known as negative entropy, anticipates the upward thrust that is evident everywhere in the plant kingdom.
This general thrust in plant life manifests in several ways which we should think of as interrelated:
Negative entropy, or storage of energy
Hierarchy
Increase of order or organization
One degree of freedom: radial symmetry
Size and growth
Self-reproduction
Evolution
But if this is the beginning of evolution, how are we to characterize the development of atoms and molecules? These too are stages of organization and as important to process as are its later stages, and they could also be called evolutionary. However, it is traditional to refer to these early stages as involution, and this word conveys the distinctive character of the left-hand side of the arc, in which process is descending, i.e., becoming more involved in matter and hence more constrained, whereas on the right-hand side it is moving up into higher forms which are more evolved and free.
In Chapter IV we described this arc with emphasis on the increasing freedom of the stages beyond the molecular. Plants at the fifth stage gain a degree of freedom which manifests not only in their growth but in their ability, through their progeny, to conquer time. Both the growth of the plant (its storage of energy, its organization) and its self-propagation through progeny result from cell division; if the cells remain together we call it growth, if they split off into a separate entity we call it progeny. But cell division is an enormously complicated process.
Complexities of the cell
It might be thought that cell division is merely an increase in size with an imposed limit which enforces division, much as water leaking from a spigot forms into drops of a certain size. But it is becoming apparent that cell division is a very complicated process. It used to be said that the cell is filled with protoplasm, but this description is as inappropriate as to say that a radio is filled with radioplasm.
The cell is the site of a vast number of complicated chemical reactions. These reactions are conducted by enzymes, specialized protein molecules that cause certain chemical reactions to occur about a million times faster than they would occur without the enzymes. The cell also includes DNA, which carries the information, and RNA, which conducts the manufacture of material for cell growth.
Escherichia coli, the bacillus which inhabits the intestine and which divides every twenty minutes, is one of the smallest life forms and the one biologists have studied most thoroughly. It is estimated to contain - in addition to 4 DNA boss molecules of a MW (molecular weight) of 2.5 billion each - about 400,000 RNA molecules (of one thousand kinds) with an average MW of 2 million, about 1 million protein molecules (of two thousand kinds) with an average MW of 40,000, plus 500 million smaller organic molecules (of seven hundred kinds) with an average MW of three hundred.
Such descriptions are, of course, mind-boggling; but to keep a sense of proportion, we should at least recognize that this is of a different order of organization from that of even the most complex molecule per se. In fact, if we were to liken a molecule to an automobile (both are combinations of different parts), the cell would be comparable to a manufacturing plant, a vast organization of men, machines and computers.
Multicellular organization
Staggering as is this achievement, it is only the beginning of the organization stage of process, i.e., the plant kingdom. The plant is able to organize a multicellular entity - say a tree - which contains trillions of cells whose cooperative interaction is essential to the organism as a whole.
How are we to deal with this kingdom? Should we turn to the biochemist, who has discovered the structure of DNA and the steps by which it conducts the internal chemistry of the cell? If we do no more than that, we will miss something which the lady who says, "My roses are happy today," has not lost touch with - the existence of the plant as a living creature. We must recognize that science can tell us of only a small part of the total picture, for the miracle of plant growth entails the development of the organization principle, and that goes far beyond the chemistry of cell division. This principle is able to guide the growth of the plant, which means that there is something which organizes the plant and is a fellow creature.
I have suggested that DNA with its possibly superconductive core, its coiling, its necessary inductance - together with the fact that all nearby cells have this same DNA - could broadcast at frequencies in the lower part of the infrared, not only to monitor the activity within its own cell, but to coordinate growth steps of neighboring cells. While this is only a supposition, it helps to delineate the problem. Something of the nature of radio broadcast is necessary to account for the coordination of cell growth.
Other principles
Meanwhile, let us proceed and keep our eyes open. Certain self-evident facts about plants, passed over by biologists, are important, such as the fact that the tree is not all alive: its wooden trunk is built up year by year by the cambium layer, which is the only living part of the trunk. Similarly, while the leaves are alive, the branches that support them are not. Thus the life of the tree is a thin surface film which covers the trunk and branches, in contrast to the animal, whose life penetrates its whole volume. This distinction, like the difference in their degrees of freedom, helps to give us an overall view of the difference between plants and animals.
Another important fact is that it is the growth principle that makes evolution possible. Current science regards evolution as explained by a process of selection, "survival of the fittest." But it does not tell us how something arises to be selected. Selection is a cutting-off process, like getting one's income from cutting coupons, but cutting off does not explain the growth that provides something to cut off. Growth in plants traces back to cell division by which both the mature plant and its reproduction come about. This growth is what is called exponential: it not only increases, but if not checked, it increases at an increasingly rapid rate.
A single bacterium like the Escherichia coli, dividing every twenty minutes, would fill a football stadium with its progeny in twenty-four hours if conditions permitted. It is such expansion that makes it possible for selection to work in order to provide a population adapted to survive in a given milieu. This expansive tendency is a major contribution to evolution.
The fork: self-limitation
This leads to another and very subtle aspect of the plant kingdom. The multicellular organism - say, a tree - is not wholly committed to self-expansion, but has the alternative of producing progeny. In this connection, it is interesting that if we wish to make a tree bear fruit, we inhibit its growth, by pruning the roots or branches or even by banding the tree to reduce the nourishment or circulation through the cambium layer. The tree takes this signal as a warning and turns its energies to producing seed. This choice, or fork, is intrinsic to the fifth stage and is an integral part of process. We may interpret it as the inverse of that which occurs in the third - self-determination. The inverse is self-surrender. Or if we read the third stage as "taking on a center," the fifth would be giving off centers, by production of seed.
Here we have a most far-reaching application of the deductive tools provided by the arc. When we read "giving up self-expansion" as the inverse of "self-determination," we might seem to be making a mere play on words. But by realizing that the one describes the creation of a nucleus, such as makes the atom possible, and the other describes the creation of seeds, which makes reproduction possible, we can appreciate that our method highlights the most conspicuous contributions of two kingdoms.
The why of the third stage: acquiring identity
But it might be asked: how can we show that self-determination, or "having its own center," and not some other manifestation, occurs at the third stage? How is this principle imbedded in the formalism? This takes us back to the degrees of freedom and constraint as set forth in the arc:
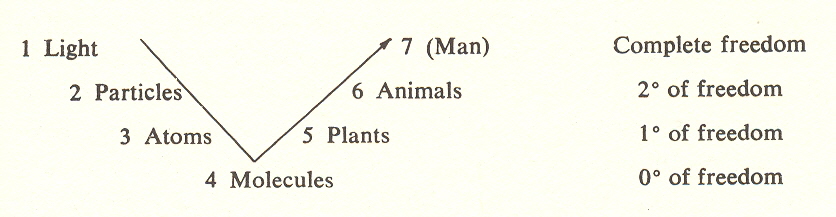
It will be recalled that the second level introduces time, and the creation of nuclear particles having substance but no identity. This lack of identity is due to the fact that there is only one constraint (time). Without another dimension, there is no way to locate or define identity. This definition is the function of the third level, which provides two constraints (dimensions) and thus a basis for definition (limitation) of the endlessness.
The atom, a typical entity, acquires an identity because it possesses a nucleus, an unchanging center, which survives the encounters that affect its periphery (its shell electrons).
The why of the fifth stage: giving up identity
In stage five we find just the reverse: the plant eventually gives up the power of unlimited self-increase, its centered negative entropy. (This sacrifice is not an evolutionary setback, for it makes possible the yet greater freedom of mobility or animation that we will take up in the next chapter.) The development of the power which leads to reproduction, and the plant kingdom, reaches its ultimate expression in the flowering plants, or angiosperms, whose perfection of the seed principle has produced the biosphere - the vegetation which covers the earth: grassland, forest, jungle.
Homogeneity of cells
One more important point about plants is that every cell of a plant is a twin brother of every other cell. The tree is a family of cells descended from one. The relationship of the cells is rooted in their DNA, so that they all have the same directive within them. Organizations of cells thus have a more cohesive origin than organizations of people. In the latter, there is always the possibility of conflict. Within a multicellular organism this cannot arise. The function which differentiates a leaf from a root, and which operates through enzymes, has its origin not in the cells, but in the needs of the organism. Apart from such functional differences, all cells of a plant are exact duplicates of one another. Their gene structure is identical. Due to cross fertilization, this identity is not present in the progeny, which have their own identity.
Substages of the plant kingdom
Having divided the atomic and molecular kingdoms into seven substages, we may now do the same for the plant kingdom.
Plant classification
But is there one plant kingdom? Oddly enough, two process philosophers, A. N. Whitehead and Oliver Reiser, who divide the great chain of being into about seven stages, agree in assigning unicellular and multicellular organisms to different kingdoms, not just different substages. Their attitude seems to gain support from the kingdoms already covered: protons and electrons when organized make atoms; atoms combine to form molecules; molecules to form cells. In each case, a plurality of units at one level is a unit at the next. So why not one kingdom for cells and another for multicellular organisms?
My answer to these philosophers is that cells introduce a higher type of organization than is involved in atoms and molecules. This new principle, based on cell division or self-multiplication, makes it possible for a single cell to create the multicellular organism out of itself. Cells are not like bricks of a house, distinct from the organism they create; they are not brought together as are atoms to make a molecule. Thus the plant kingdom, which develops the power of organization and does so through cell division, cannot be divided into two kingdoms.
Biologists go to the other extreme and do not even regard the distinction between uni- and multicellular as a basis for classification. They divide algae (seaweed) into seven or eight grand divisions irrespective of the uni/multi distinction. Some algae (euglena and diatoms) are unicellular, and others include both unicellular and multicellular forms.
As may be seen in the accompanying chart, there is considerable difference in systems of classification. The 1942 division shown uses seven phyla (the phylum is a major division) on algae, three on fungi, one on moss, and then puts all higher plants in one phylum! This has always seemed absurd to me, and I am glad to see that Bold, from whose book The Plant Kingdom the table is taken, adds nine major divisions to cover higher plants. But while the twenty-four divisions so obtained are an improvement, they are not much help in our problem of levels of organization.
Here it is interesting to note, however, that when Bold discusses the development of the plant kingdom, his chapters follow closely the division we propose concerning that development.
Our discussion of the diversity of the plant kingdom has included reference to such groups of organisms as algae, fungi, mosses. . . gymnosperms and angiosperms. Could we have started as well at the end of the series or perhaps with any intermediate group and proceeded in a different order? The order of our study is indeed significant for it reflects a series of organisms of increasing complexity.*
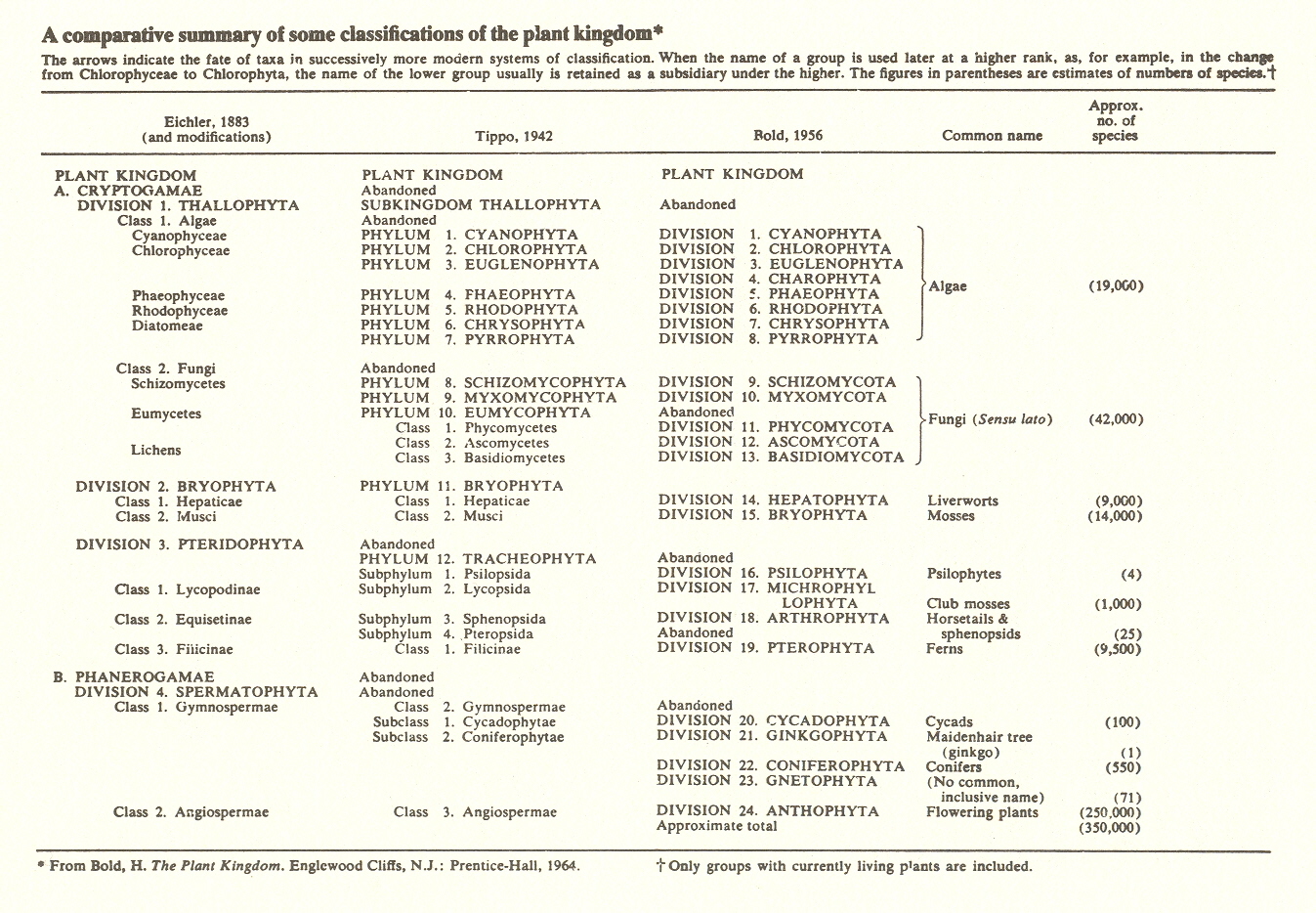
Clearly, Bold feels the importance of the levels of increasing complexity which we are stressing, so let us proceed.
First substage
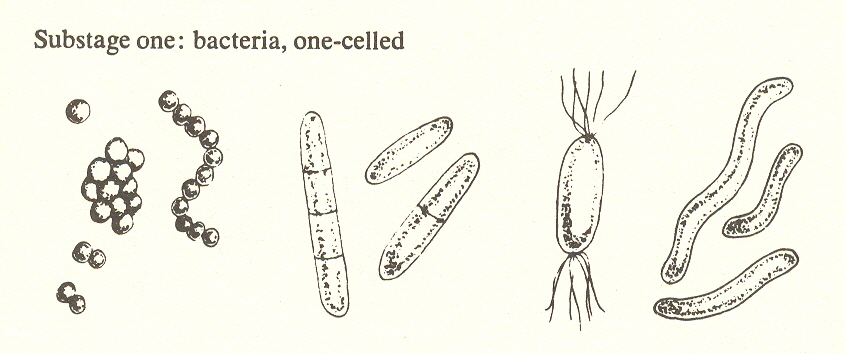
Despite the fact that biologists attach so little importance to the uni/multicellular distinction, we must stress that unicellular plants are the first step toward the complexity of which Bold speaks. By definition, they cannot have cell differentiation, and thus they qualify as first in order of complexity. Here we have bacteria and diatoms, all microscopic.
Second substage
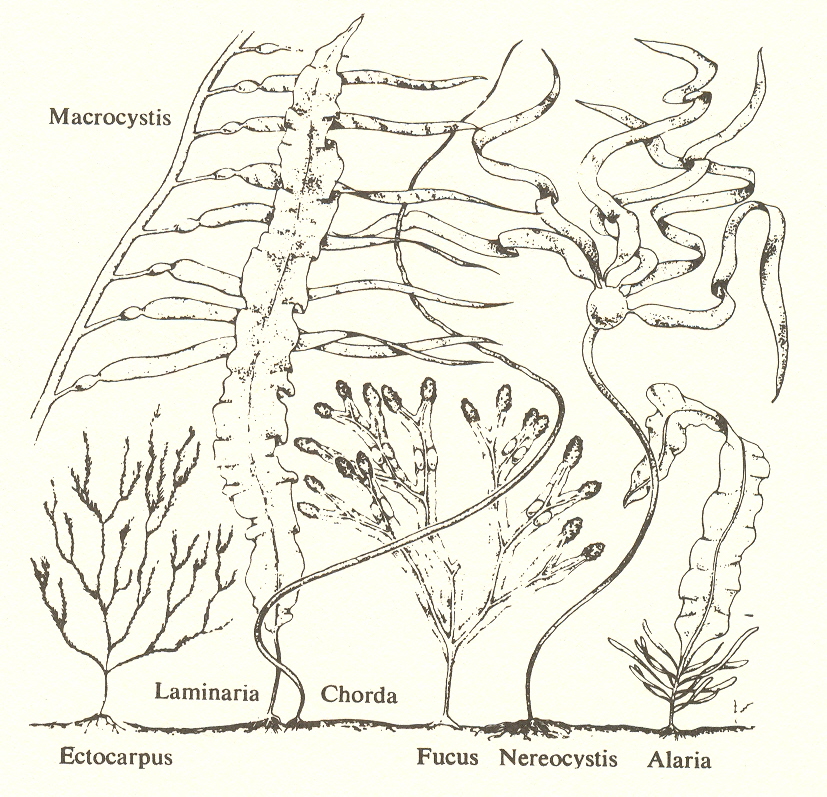
The conspicuous development here is a kind of binding that gives rise to great size. Millions of cells adhere together to form a tissue - a plant as we know it, even though a simple one. Here we have seaweed, including the giant kelp which grows over a hundred feet long. Cell differentiation also has begun; there are stem, leaves, and the holdfast cells by which the plant is attached to the ocean floor. We also find the differentiation of cells required for sex instead of two similar cells conjugating, egg and sperm are differentiated, differing in size and number. And the sperm cells have to swim to the egg.
Substage two: algae, many-celled (from Villee, C. A., Biology, Philadelphia: W. B. Saunders Co., 1972)
Third substage
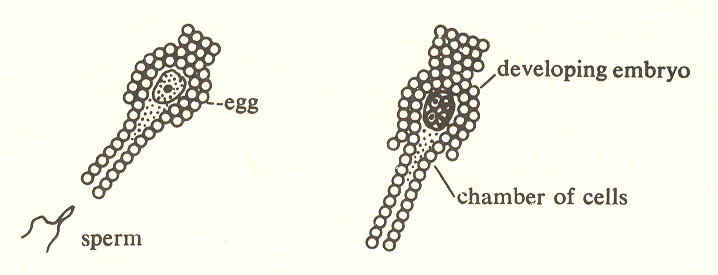
All biologists agree that the next development is the mosses and liverworts, the first land plants. These are Embryophyta ("bearing embryos"). They are the first plants to differentiate the embryo from the rest of the plant by forming a chamber of cells, with the young plant (itself a multicellular organism) within it.
This is a bull's eye for process theory, because here we have identity expressed in terms of reproduction. The embryo is given an identity.
All higher plants are Embryophyta, which is to be expected in view of the cumulative nature of process, each stage or substage incorporating what has gone before. So, in order to limit the third substage to the first plants to develop an embryo, we should designate it as Bryophyta. "Bryos" means moss and has no etymological connection with "embryo." I have used the word Embryophyta in the grid because it is more descriptive of the principle which emerges here.
Substage four: psilophytes, vascular tissue (from Villee, C. A., Biology, Philadelphia: W. B. Saunders, 1972)
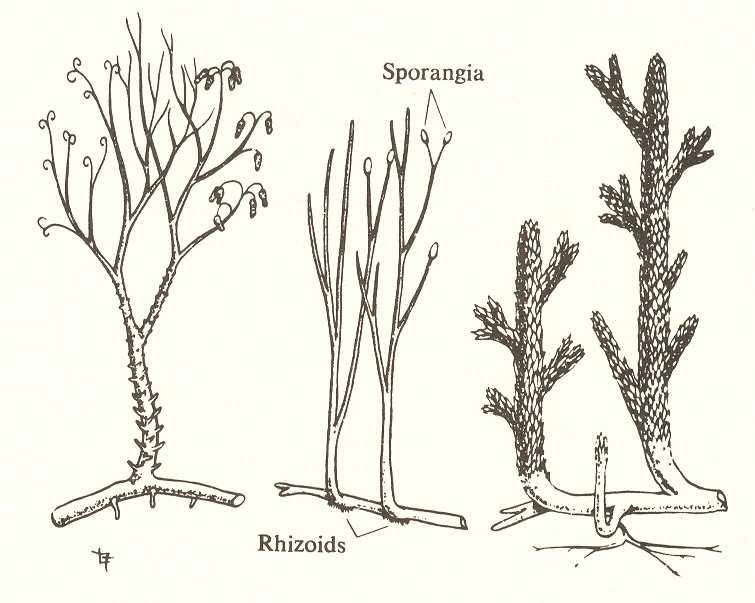
Fourth substage
The next development in plants is vascular tissue, the xylem and phloem which conduct fluid and food through the plant and thus make possible an increase in size. This substage would include psilophytes and club mosses.
Substage five: calamites (horsetails), segmented vascular tissue (Villee)
Fifth substage
At this point there is a most important development insufficiently stressed by Bold: segmented vascular tissue, which is the innovation that makes the great difference in size between club mosses and trees. The plants that exhibit this innovation are the Equisetineae or horsetails, called Arthrophyta by Bold and Calamites or Sphenopsida by others. Currently, these are not important plants, but they introduce the production of supportive tissue, and in Carboniferous times reached a height of ninety feet. The strength of modern trees is due to segmented tissue in the form of wood. The correspondence here is that the fifth substage in general is chain-like. Examples: the chains of cells in all plants, the chains of molecular units in polymers.
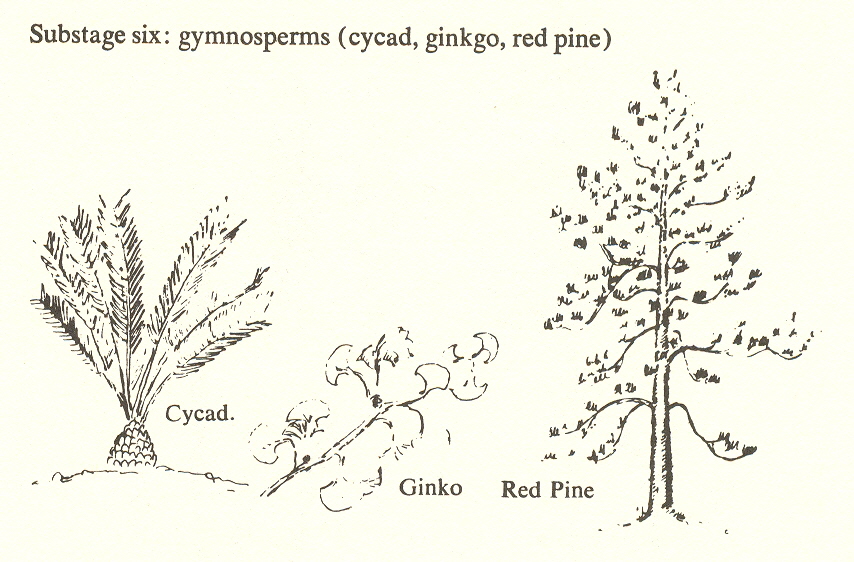
Substage six: gymnosperms (cycad, ginko, red pine)
Sixth substage
We now come to the plants known as gymnosperms, which include the conifers - pine, spruce, cypress, hemlock, etc. - and which represent, we believe, the sixth substage of the plant kingdom. These are the first of the seed-bearing plants. (The term "gymnosperm" means that the seeds of these plants are naked, i.e., not enclosed in an ovary.) And the factor of mobility which we expect at the sixth substage is exhibited in the mobility of the seeds.
Digression on purposiveness
It would appear that, besides growth and reproduction, there are two problems: the distribution of the plant over wide areas and the crossing of genes with other plants of like species. The former is achieved by the transport of spores or seeds, and the latter by having the eggs of a given plant fertilized by sperm from a separate plant, as when pollen is carried from one flower to another.
But in the more primitive plants, the sperm cannot reach a different plant; it has to swim over the surface of the gametophyte (as in mosses and ferns) . One might think that these would be placed close together for the more certain achievement of fertilization. But no, in moss the sperm must swim to the top of a long pointed archegonium, and in the fern the sperm is produced at the opposite end of the gametophyte and has to swim its entire length. Why? This question is not answered or even raised in the texts I've consulted, but I think it is important because it illustrates the purposiveness which we have referred to earlier as an important "category of explanation" (Whitehead's phrase), and one that has a role throughout evolution.
To repeat the argument of Chapter II, action
is a recognized measure of science, the integral of energy over time (Action = E x T). It was given importance by Planck's discovery that action occurs in wholes or quanta that are as fundamental as the proton and electron, if not more so. This wholeness, in conjunction with the fact that action includes mass, length, and time as "parts," enables us to say that M, L, and T are derived from action, the whole. Purpose, which of necessity cannot exist in the part, can however be considered to exist for the whole. Indeed, it is synonymous with the function which the whole, and only the whole, can exhibit. (See end of Chapter II.)
In the plant kingdom we encounter a reassertion of this purposiveness that has been dormant in the atom and molecule. The sperm, having the initiatory function in reproduction, can be correlated to the photon, or to first cause, in the life cycle. Like the photon, it is action before its manifestation in matter. Thus the journey of the sperm to the egg is the purposive stage of process; and I believe the separation of sperm and egg is an evolutionary device to ensure that purposiveness rather than accident start each new generation. It would be perfectly simple to have the sperm produced very close to the egg, or even in the same compartment, but then accident would provide fertilization with no intention. It is necessary to have a distance between the sperm and the egg in order for the teleological factor to manifest.
How can this be done without reducing the likelihood of fertilization? If it did reduce that likelihood, the progeny would be reduced and the species would suffer and perhaps die out. The answer solves another question that biologists neglect: why are so many sperm produced that one egg may be fertilized? It is by an enormous increase in the number of sperm, and a vast decrease in the likelihood of any given sperm reaching the egg, that the teleological factor is enabled to assert itself.
It could be objected that this is merely a question of the survival of the fittest sperm. But we must recognize that since the sperm is only one cell, its survival cannot be based on the multicellular structure encoded in its DNA, because this structure is as yet unmanifest, and therefore not helpful. The sperm's survival depends, rather, on its liveliness, or on its persistence in moving toward the egg, i.e., on its purposiveness.
Seventh substage
The seed-bearing plants known as angiosperms constitute the seventh substage of the plant kingdom. The angiosperm ("vessel-seed") is distinguished from the gymnosperm ("naked-seed") in that the seed has a covering. This is the crowning touch that gives the angiosperms their dominance of the plant kingdom. Of the 350,000 species of all plants, 250,000, or 5/7, are angiosperms. Of land plants, they represent an even greater portion. All flowers, grasses, and hardwood trees are angiosperms. The covering for the seed makes possible a variety of devices which aid in seed transport. Such are thistles, burrs, maple samaras which auto-rotate. So too, nuts, grains, and fruit which animals and birds transport and thereby distribute. Man's main food supply is either directly dependent on such seed covering as in the case of wheat, corn, grains, and vegetables, or indirectly in the case of grazing animals.
Tissue layers
While our division of the plant kingdom is less clear-cut than that of the atomic kingdom, where the relative simplicity of the periodic table gives us a clear structural example of sevenfoldness, or of the molecular kingdom where the powers of binding, identity, combination, growth, mobility, and dominion find expression in the division of molecules supplied by Dr. Price, nevertheless the proposed division at least furnishes an overall survey of the plant kingdom. Even if not ultimately correct, it can serve as a basis for a more penetrating examination of the principles of organization.
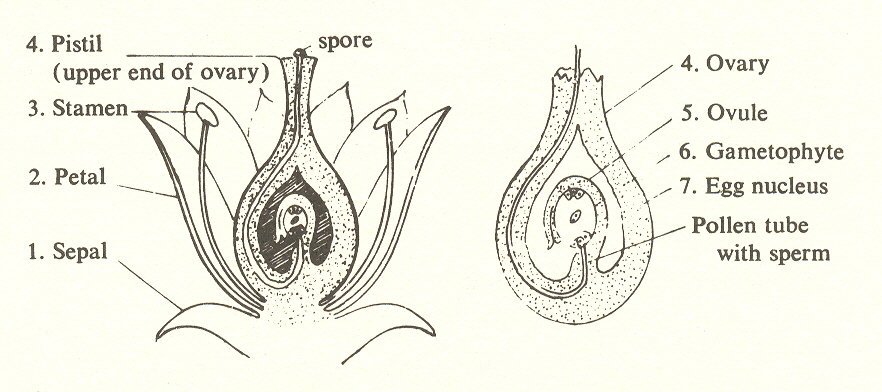
What, then, can we learn from this division of the plant kingdom into seven substages? A hint comes from the fact that the angiosperms, or flowering plants, show in cross section seven layers of tissue:
Like the seventh-row atoms, the flowering plant has seven layers or "shells".
The flower is surrounded by an outer layer of sepals (1) which enclose a second layer of more conspicuous petals (2), within which are the stamens (3), source of the pollen, and within that the pistil, which is at the upper end of the ovary (4). Within the ovary are a number of ovules (5), within which is the gametophyte (egg-bearing) (6). Inside of this are eight nuclei, one of which becomes the egg (7), and is fertilized by a nucleus from the pollen microspore. (The process is complemented by a second sperm nucleus in the pollen microspore which fertilizes two other eggs to provide the coating for the seed.
Without laboring the possibility that this is a valid seven-ness, let us note that there are resemblances to the atom, for the angiosperm in having seven layers is analogous to the complex atoms with seven shells of electrons. There might even be a correspondence in the two nuclei of the microspore and eight of the gametophyte with the 2- and 8-shells of electrons. But if this is true of the angiosperms, how about the rest of the plants? It is a project that would require microscopic examination. However, inasmuch as the sixth-substage gymnosperms ("naked-seeds") have by definition one less layer than angiospermsand since unicellular plants have by definition only one layer-we could say that insofar as evidence is available a correspondence between the number of layers and the substages is borne out.
The number of layers of tissue may, in any case, be a manifestation of a deductive principle and hence a subject for study, and possibly a basis for classification. Recall the Pauli exclusion principle, which so precisely accounts for the seven possible shells of atoms. The layers have an additional significance in that they also represent stages in the life cycle of plants. Thus the plant starts as a seed, it grows, it blossoms, it is fertilized, it fruits, and finally the fruit decays, leaving the seed which still contains the germ plus a coating of starch that provides nourishment for the infant plant.
Fungi and retrogressive stages
Fungi, an important division of plants which includes molds, lichens, and mushrooms, are not included in the scheme described so far. This omission would be a serious objection to the sevenfold division of the plant kingdom were it not that the other kingdoms include entities that do not follow the pattern. What is unique about fungi is that they have no chlorophyll and do not engage in photosynthesis, which means that they do not, like all other plants, draw energy from sunlight and store order against the flow of entropy; they are not moving up the arc.
Similarly, there are atoms which do not naturally combine with other atoms. These are the noble gases: helium, argon, neon, krypton, xenon, and radon. As for particles, there is the so-called antimatter, which reverts to radiation. The animal kingdom too has its "won't play" entities, the tunicates, sessile animals, which are actually Chordata, the highest subdivision, but resign their opportunity to use mobility and revert to a sponge-like existence.
To complete the scheme, we would expect molecular entities that remain in limbo and do not "progress." This category is actually most obvious of all: the molecules that constitute minerals, which take no active part in evolution.
The addition of these "whiskers" to the arc does not change the sevenfold nature of process. It does indicate the continual presence of option in that, at each stage, there is a choice which, if not made correctly, results in a renunciation of evolution.